 Science and Technology
Science and Technology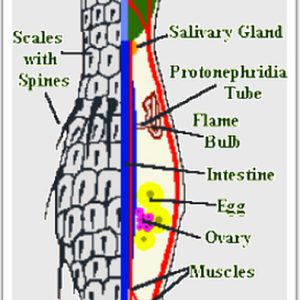 Gastrotrich Morphology
Gastrotrich Morphology 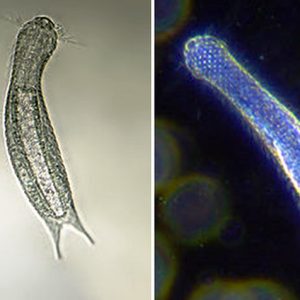 Gastrotrichs
Gastrotrichs
501-918-3025
calsfoundation@cals.org
calsfoundation@cals.org
The Phylum Gastrotricha (commonly called “hairybacks”) comprises about 800 species of marine, brackish, and freshwater microscopic invertebrates. Twelve genera and fewer than 100 species of freshwater gastrotrichs are known from North America. However, in North America, perhaps 75 to 90 percent of the probable diversity of freshwater gastrotrichs species are undescribed.
Gastrotrichs are widely distributed cosmopolitan organisms that are divided into two orders: the Macrodasyida (with nine families), which, except for two freshwater species (Marinellina flagellata and Redudasys fornerise), are marine, and the Chaetonotida (seven families), some of which are marine, estuarine, and semi-terrestrial forms, while others are primarily found in freshwater. The Chaetonotida can be further broken down into two suborders: the Multitubulatina (Nesodasys) and the Paucitublatina (Chaetonotus, Dasydytes, and Lepidodermella). Of these, the genus Chaetonotus typically dominates gastrotrich faunas everywhere in fresh water, both in terms of numbers of species and in numbers of individuals. The phylum was named in 1865 by Russian zoologist Ilya Ilyich (Élie) Metchnikoff (1845‒1916).
This group of invertebrates has been called an enigmatic protostome phylum, as scientists are still uncertain about how it fits into the Tree of Life and the relationship of gastrotrichs to other phyla has proven puzzling.
Seven major characteristics of the phylum include: (1) bilateral, triploblastic, a-segmented, and pseudocoelomate bodies; (2) elongate body or a body shaped like a bowling pin, and microscopic (0.06 to 3.0 mm or 0.002 to 0.118 in.) in body length; (3) dual gland adhesive tubes (glands); (4) complete gut; (5) smooth bi-layered (unmolted) cuticle or with scales/spine-like elaborations with exocuticle covering entire body including all cilia; (6) direct development; and (7) complex reproductive organs, either hermaphroditic, parthenogenetic, or both.
These organisms are mostly benthic and live within the periphyton, the layer of tiny organisms and detritus that is found on the seabed and the beds of other water bodies. The majority live on and in the interstitial spaces between particles of sediment or on other submerged surfaces, but a few species are terrestrial and live on land in the film of water surrounding grains of soil. They also inhabit the surfaces of aquatic plants and other submerged objects. In addition, they are found in stagnant pools and anaerobic mud, where they thrive even in the presence of hydrogen sulfide gas. When pools they inhabit dry up, they can survive periods of desiccation as eggs, and some species are capable of forming cysts in harsh environmental conditions, including those that are anoxic.
Gastrotrichs are colorless organisms that have a simple body plan consisting of a cuticle with a head and trunk region, a brain with sensory organs, a muscular pharynx, and a trunk with a simple gut and reproductive organs. Most gastrotrichs have a neck-like constriction behind the head. The pharynx opens into a cylindroidal intestine, which is lined with glandular and digestive cells. The anus is located on the ventral surface close to the posterior of the body. In some species, there are pores in the pharynx opening to the ventral surface; these contain valves that may allow egestion of any excess water swallowed while feeding. They also have adhesive glands with which they can anchor themselves to the substrate and cilia in which they move around. Usually present is a ring of motile cilia that lines the head dorsally whereas other ciliary elaborations such as pits and tentacles occur on the sides. The trunk ends in a rounded, forked caudum or bi-lobed tail that possesses adhesive tubes.
Gastrotrichs form an integral part of the benthic community in marine and freshwater environments. They feed on basically any organic material they find, whether it is alive or dead. More specifically, they are detritivores and are microphagous, sucking organic material, bacteria, diatoms, and small protists into their mouths by pumping food items via the muscular action of the pharynx into the gut. The mouth is surrounded by a ring of stiff cilia that may assist in capturing food items. The intestine is a straight tube ending in the anus, where waste products are passed out.
Reported predators of gastrotrichs include heliozoan and sarcodine amoebae, cnidarians, turbellarians, and tanypodine midges, but several other benthic predators presumably feed on them.
Reproduction and reproductive behavior of gastrotrichs is complex but has been rarely studied. Most gastrotrichs are hermaphroditic or a combination of hermaphroditic and parthenogenetic. Many macrodasiyds are simultaneous hermaphrodites possessing both male and female sex organs, including the marine species producing eggs that develop directly into miniature adults. The freshwater chaetotonid species are parthenogenetic, producing unfertilized eggs, and at least one species (Urodasys viviparus) is viviparous. The male system is composed of single or paired testes with their associated sperm ducts that lead to single or paired ventral gonopores. In addition, a specialized caudal organ that functions in the transfer of sperm is present. The female reproductive system includes one or two ovaries in the posterior portion of the body. The eggs of all gastrotrichs undergo direct development and hatch into miniature versions of the adult. As is the case in most protostomes, development of the embryo is determinate, with each cell intended to become a specific part of the organism’s body. The juveniles typically reach sexual maturity rapidly in about three days and have lifespans of a few weeks.
Their excretory system is primitive, but nitrogenous wastes are likely excreted through the body wall, as part of respiration, and the protonephridia are believed to function mainly in osmoregulation. There are no discrete respiratory or circulatory organs. The nervous system is relatively simple. The brain consists of two ganglia, one on either side of the pharynx, connected by a commissure. The primary sensory organs are bristles and ciliated tufts of the body surface that function as mechanoreceptors. There are also ciliated pits on the head, simple ciliary photoreceptors, and fleshy appendages that act as chemoreceptors.
Gastrotrichs demonstrate a phenomenon termed “eutely,” in which each species, as adults, has an invariant genetically fixed number of cells. Cell division terminates at the end of embryonic development, and further growth is due solely to enlargement of cells.
For those interested in collecting gastrotrichs, taking samples of sediments (small diameter cores) or vegetation using very fine mesh is recommended. For quantitative work on sediment-dwelling species, small diameter (2–5 cm) cores are preferable. For preservation, the organisms can be narcotized with one percent MgCl2 for ten minutes, followed by fixation in ten percent formalin with Rose Bengal. Sometimes gastrotrichs can be handpicked and counted under a stereoscopic (dissecting) microscope, but this procedure can be quite tedious. Density gradient centrifugation can be useful in extracting gastrotrichs from sediment. A modified Baermann funnel has been useful for extracting chaetonotidans, but not dasydytids, from pond sediments. For serious taxonomic work, videomicroscopy of living gastrotrichs can provide better permanent documentation of species characteristics rather than using killed specimens or microscopic slides of voucher material.
While there are over 800 species of gastrotrichs described worldwide, little is known about this phylum in Arkansas. One early study of gastrotrichs from twenty-three sites in Washington County was conducted in 1937 for a master’s thesis at the University of Arkansas (UA) in Fayetteville (Washington County). It reported eighty-one species and six genera within a single family. In a study of organisms associated with the shell (carapace and plastron) of two species of aquatic turtles, Chaetonotus sp. was reported from those collected in Montgomery County. A modern survey of the gastrotrichs of the state is sorely needed.
For additional information:
Balsamo, M., and M. Antonio Todaro. “Gastrotricha.” In Freshwater Meiofauna: Biology and Ecology, edited by S. D. Rundle, A. L. Robertson, and J. M. Schmid-Araya. Leiden: Backhuys Publishers, 2002.
Balsamo, M. J., L. d’Hondt, J. Kisielewski, and L. Pierboni. “Global Diversity of Gastrotrichs (Gastrotricha) in Freshwater.” Hydrobiologia 595 (2008): 85‒91.
Brunson, R. B. “Gastrotricha.” In Fresh-Water Biology, 2nd ed., edited by W. T. Edmondson. New York: John Wiley and Sons, 1959.
———. “The Life History and Ecology of Two North American Gastrotrichs.” Transactions of the American Microscopical Society 68 (1949): 1–20.
Brusca, Richard C., W. Moore, and S. M. Shuster. Invertebrates. 3rd ed. Sunderland, MA: Sinauer Associates, 2016.
Davis, Kenneth B. “The Gastrotricha (Chaetonotoidea) of Washington County, Arkansas.” MA thesis, University of Arkansas, Fayetteville, 1937.
Davison, D. B. “A New Species of Gastrotrichan-Chaetonotus robustus, New Species.” American Museum Novitates 972 (1938): l‒6.
Dunn, G. W., C. Giribet, G. D. Edgecombe, and A. Hejnol. “Animal Phylogeny and its Evolutionary Implications.” Annual Revues of Ecology and Evolution 45 (2014): 371‒395.
Hatch, Melville H. “Notes on Two Species of Gastrotricha from Washington.” American Midland Naturalist 21 (1939): 257–258.
Hawkes, J. A. “A Taxonomic Study of Gastrotricha from the Southern Appalachian Mountain Region.” MS thesis, Wake Forest University, 1965.
Hochberg, Rick, and M. K. Litvaitis. “Macrodasyida (Gastrotricha): A Cladistic Analysis of Morphology.” Invertebrate Biology 120 (2001): 124‒135.
———. “Phylogeny of Gastrotricha: A Morphology-Based Framework of Gastrotrich Relationships.” Biological Bulletin 198 (2000): 299–305.
Jacobs, Madeleine. “Watch Out for Water Bears and Mud Dragons While at the Beach.” Northwest Arkansas Times, July 10, 1983.
Kånneby, Tobias, and Rick Hochberg. “Phylum Gastrotricha.” In Thorp and Covich’s Freshwater Invertebrates, 4th ed. New York: Academic Press, 2010.
Kieneke, A., O. Riemann, and W. H. Ahlrichs. “Novel Implications for the Basal Internal Relationships of Gastrotricha Revealed by an Analysis of Morphological Characters.” Zoological Science 37 (2008): 429‒460.
Manylov, O. G., N. S. Vladychenskaya, I. A. Milyutina, O. S. Kedrova, N. P. Korokhov, G. A. Dvoryanchikov, V. V. Aleshin, and N. B. Petrov. “Analysis of 18S rRNA Gene Sequences Suggests Significant Molecular Differences between Macrodasyida and Chaetonotida (Gastrotricha).” Molecular Phylogenetics and Evolution 30 (2004): 850‒854.
Pennak, Robert W. Freshwater Invertebrates of the United States, 2nd ed. New York: John Wiley and Sons, 1978.
Ruppert, E. E. “Gastrotricha.” In Introduction to the Study of Meiofauna, edited by R. P. Higgins and H. Thiel. Washington DC: Smithsonian Institution Press, 1988.
Strayer, David L., William D. Hummon, and Rick Hochberg. “Gastrotricha.” Ecology and Classification of North American Freshwater Invertebrates. Cary Institute of Ecosystem Studies, 2010. Online at: https://www.caryinstitute.org/sites/default/files/public/reprints/thorp_covich_gastrotrichs_2010.pdf (accessed January 10, 2020).
Todaro, M. Antonio, M. Dal Zotto, U. Jondelius, Rick Hochberg, W. D. Hummon, Tobias Kånneby, C. E. F. Rocha. “Gastrotricha: A Marine Sister for a Freshwater Puzzle.” PLoS ONE 7 (2012): e31740
Tumlison, Renn, and Sharon Clark. “Microorganisms Associated with the Carapace and Plastron of Aquatic Turtles (Pseudemys concinna and Trachemys scripta) in Southwestern Arkansas.” Proceedings of the Arkansas Academy of Science 50 (1996): 148‒152.
Weiss, M. J. “Widespread Hermaphroditism in Freshwater Gastrotrichs.” Invertebrate Biology 120 (2001): 308‒341.
Zhang, Z.-Q. “Animal Biodiversity: An Introduction to Higher-Level Classification and Taxonomic Richness.” Zootaxa 3148 (2011): 7–12.
Henry W. Robison
Sherwood, Arkansas
Chris T. McAllister
Eastern Oklahoma State College





Comments
No comments on this entry yet.